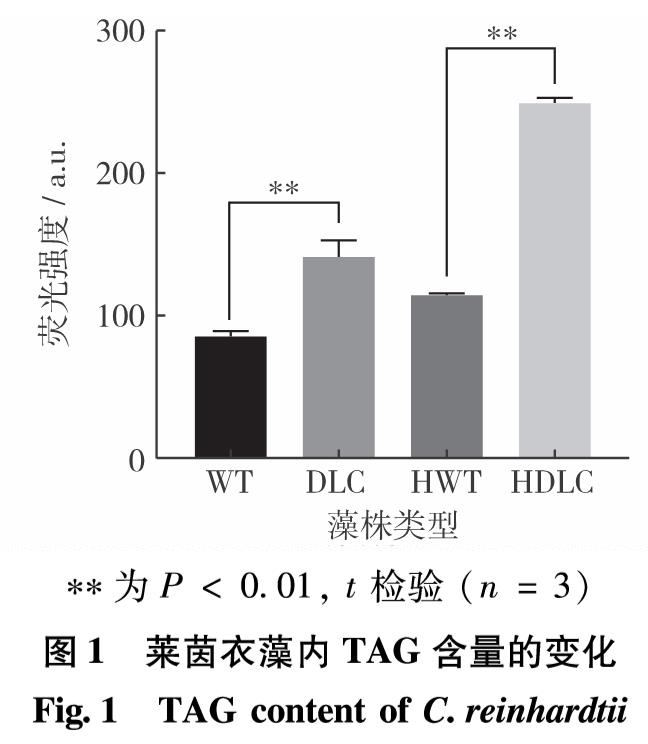
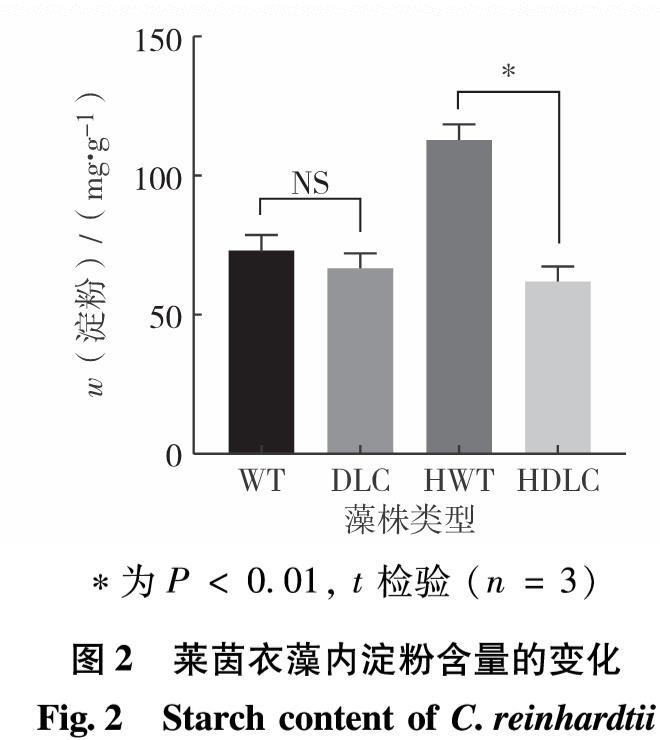
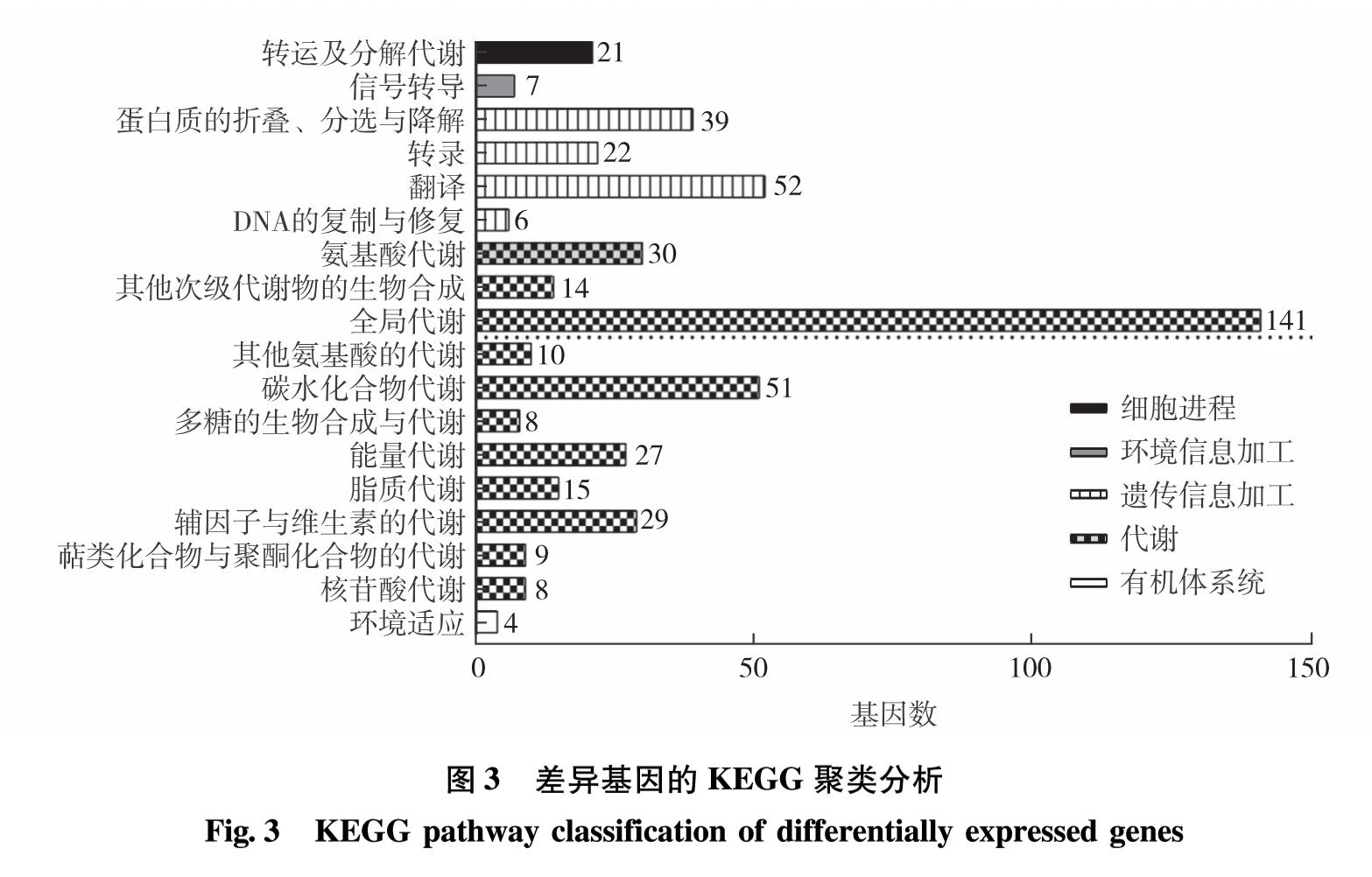
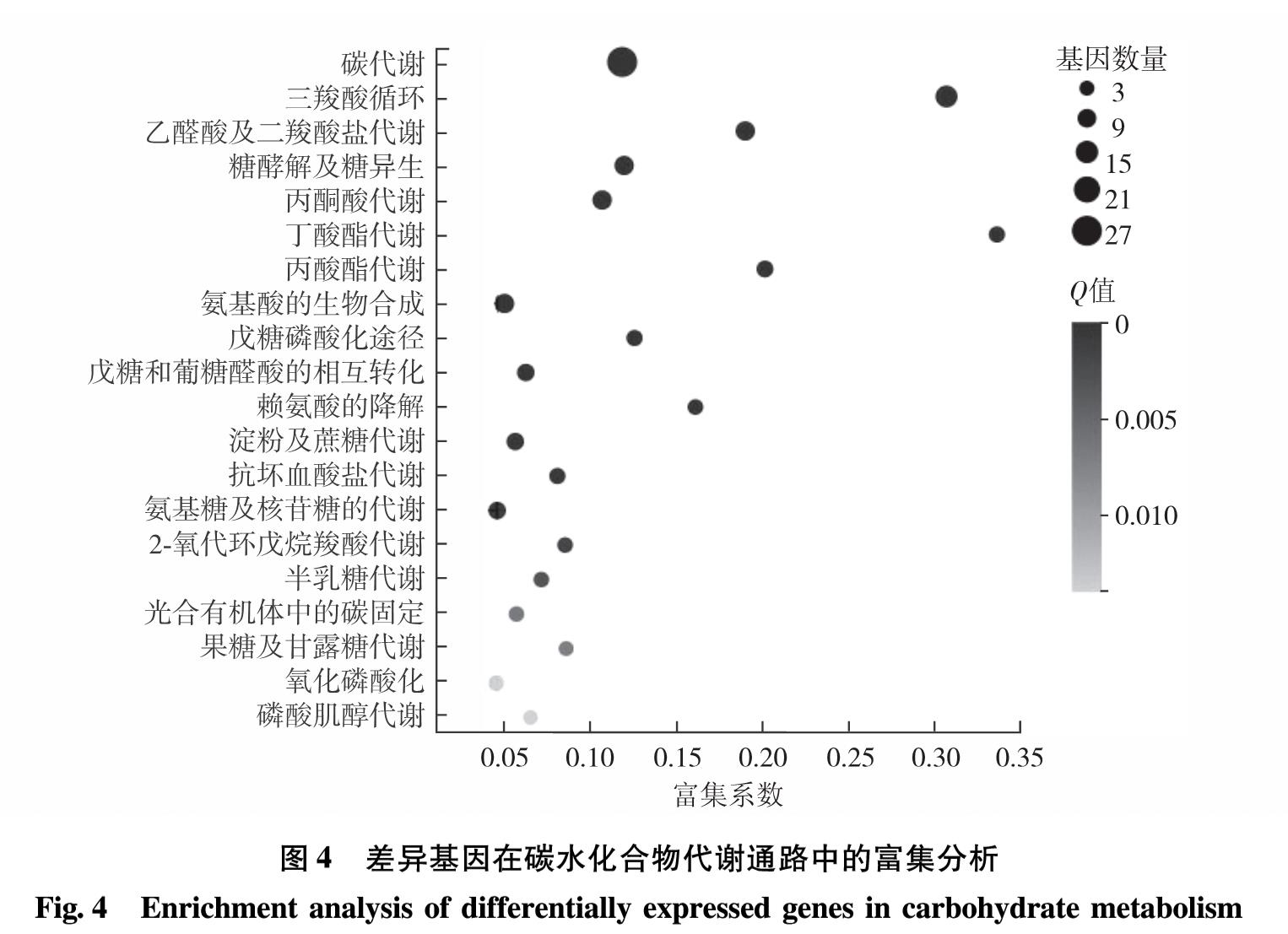
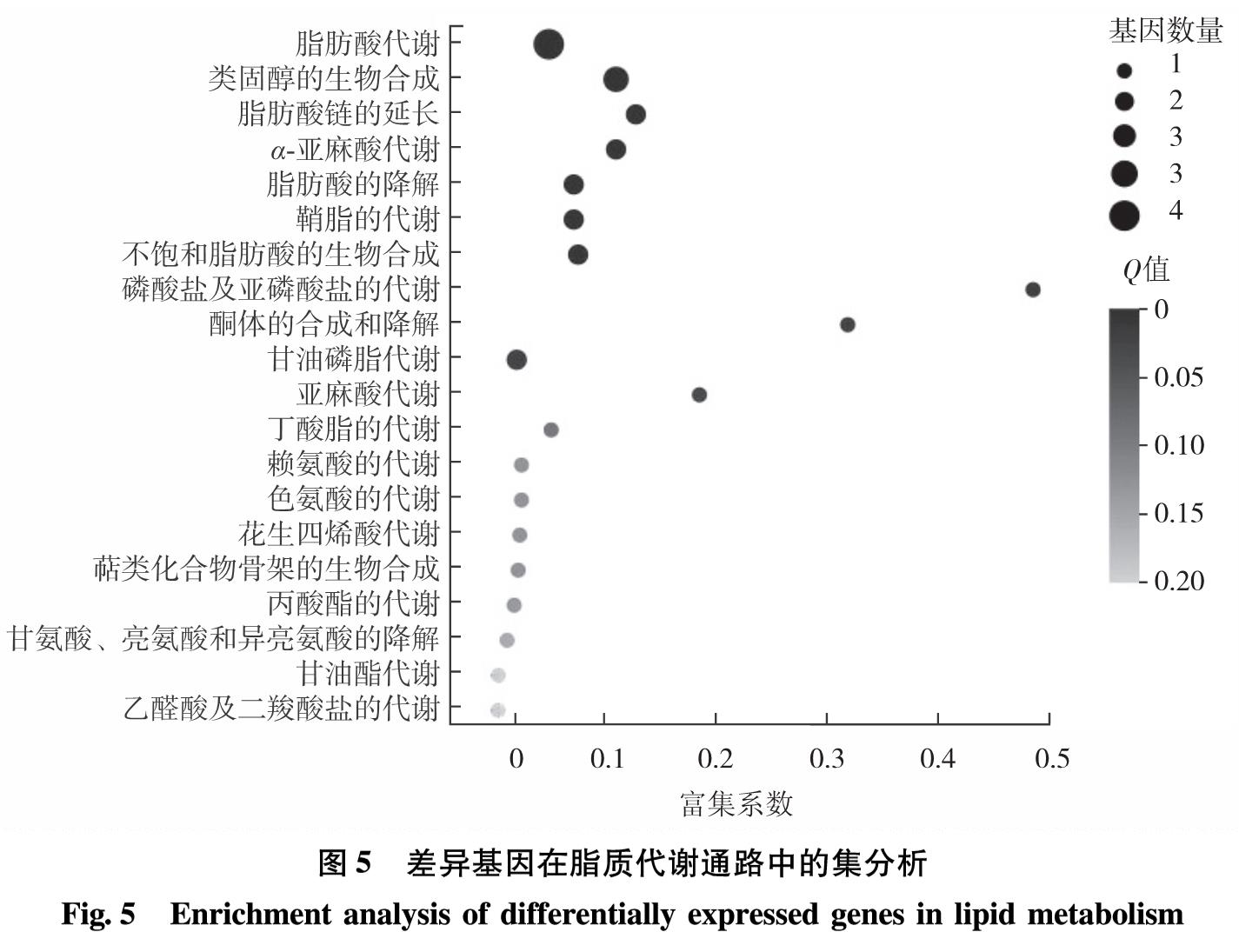
脂质合成代谢中的甘油三酯代谢、甘油磷脂代谢、脂肪酸代谢等途径与糖代谢中的糖酵解、糖异生和淀粉代谢等途径相关联,因此,若其他代谢通路含有生成三磷酸甘油、甘油、乙酰辅酶A和脂肪酸等共同中间代谢产物,则能在一定的调控条件下改变碳流的偏向,促进脂质合成代谢. ACS2和CIS1分别参与脂质合成及糖代谢中的关键反应,而DOF转录因子介导碳氮代谢中多通路的调控,则3基因的共同调控定能引起其他关键基因表达水平的变化.
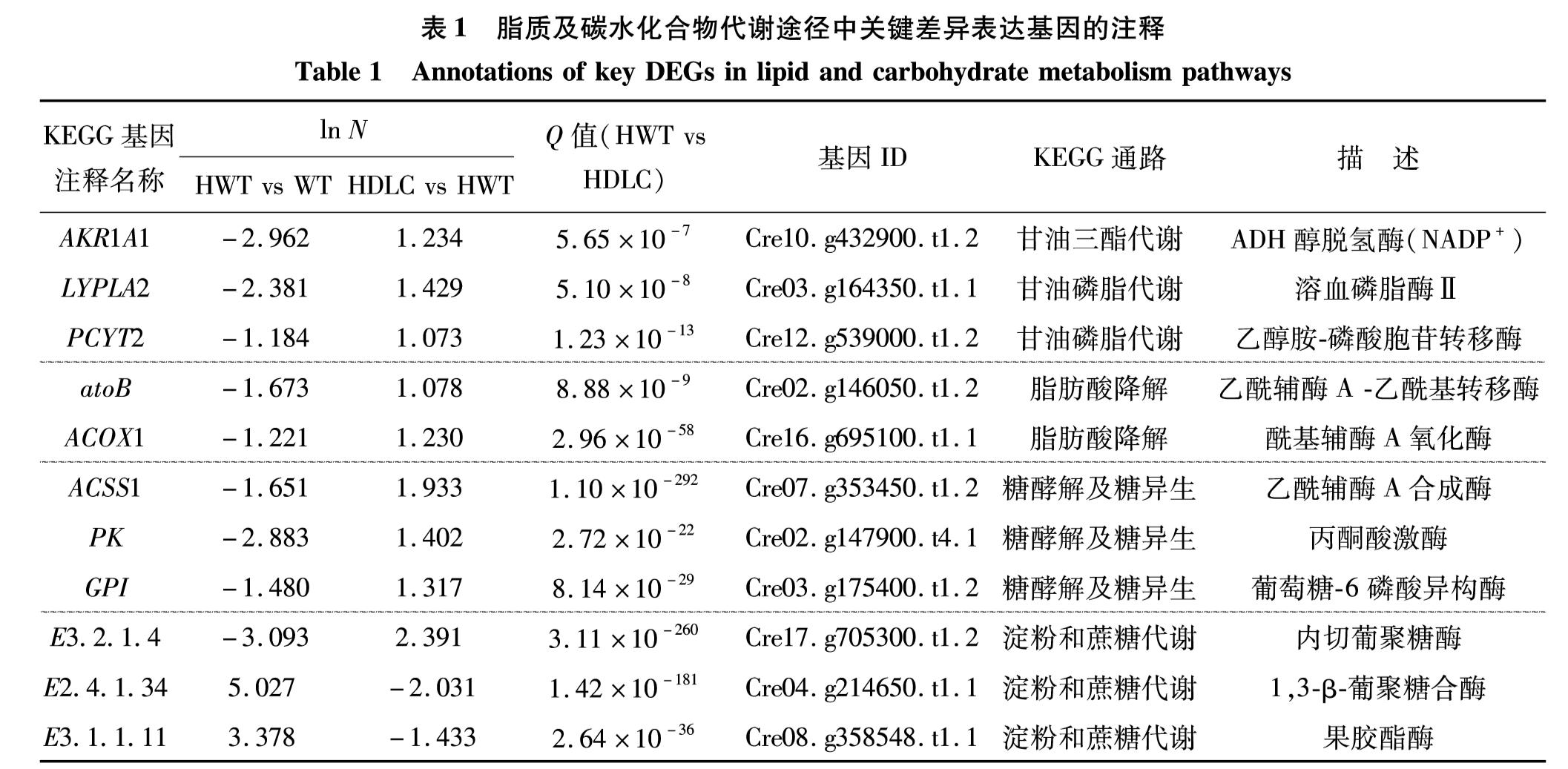
在甘油三酯代谢中, 醇脱氢酶(alcohol dehydrogenase, ADH)能将甘油氧化为D-甘油醛,再被甘油脱氢酶催化为甘油.有研究表明,编码ADH及乙酰乙酸脱羧酶等参与乙酰辅酶A代谢的基因的共同过表达导致大肠杆菌内的脂肪酸异丙基酯达203.4 mg/L[11].与WT藻细胞相比,ADH在HWT 藻细胞中的表达水平下降; 而相比于HWT藻细胞,ADH在HDLC中表达水平上调,说明3基因调控下ADH的上调可能导致甘油的增加而增强脂质合成代谢.
在甘油磷脂代谢中,三磷酸甘油是重要的中间产物.相较HDLC藻细胞中溶血磷脂酶Ⅱ(lysophospholipase Ⅱ, LYPLA2)及乙醇胺-磷酸胞苷转移酶2(phosphate cytidylyltransterase 2, PCYT2)的表达量增加,LYPLA2以2-酰基甘油-3-磷酸胆碱为底物,催化生成甘油-3-磷酸胆碱,随后被转化为三磷酸甘油; 而PCYT2将磷酸乙醇胺转化为CDP-乙醇胺,产物经接连反应后被转化为三磷酸甘油.WEPY等[12]发现LYPLA1和LYPLA2共同调控神经细胞脂质代谢的平衡,任何一方的活性缺失均会导致溶血磷脂的增加; PAVLOVIC等[13]则发现pcyt2的沉默导致小鼠胚胎细胞内磷脂酰乙醇胺合成的提高,使得膜脂增加.因此,热激后3基因的调控下LP2、 PCYT2的表达上调可能抑制膜脂的合成,使得脂质的生物合成进程偏向三酰甘油的合成.
在脂肪酸降解途径中,乙酰辅酶A是由脂肪酸β-氧化和糖酵解终产物丙酮酸经氧化脱羧产生的,在有机体中的许多代谢过程中发挥着关键的作用.β-氧化的连续周期反应过程中,每一步反应中长链脂肪酸脱去含两个碳原子的碎片而被氧化为逐缩短碳链的脂酰辅酶A,从而产生乙酰辅酶A[14].相比HWT藻细胞,HDLC藻细胞中酰基辅酶A氧化酶1(acyl-CoA oxidase, ACOX1)和乙酰辅酶A乙酰基转移酶(acyl-CoA acyltransferase, ACAT)的表达量上调. ACOX1参与多种碱基辅酶A的氧化反应,促进脂肪酸的氧化分解[15]; 而ACAT催化短链脂肪酸的氧化反应,生成乙酰辅酶A[16].故而两个基因的上调表达可能一定程度上促进脂肪酸的氧化分解,加快乙酰辅酶A 循环流的流动,进而提高脂肪酸合成的中间产物循环效率.
丙酮酸既是糖酵解途径的终产物,也为糖异生途径的起始反应物.作为乙酰辅酶A的合成前体,丙酮酸与脂质合成代谢有着紧密的联系.在糖酵解及糖异生代谢中,相较于HWT藻细胞,HDLC藻细胞中的葡萄糖- 6-磷酸异构酶(glucose- 6-phosphcrte isomerases, G6PI)、 丙酮酸激酶(pyruvate kinase, PK)及乙酰辅酶A合成酶(acyl-CoA synthetase, ACS)表达水平更高.G6PI催化α-D- 6-磷酸葡萄糖和β-D- 6-磷酸葡萄糖间的相互转变,及两者向β-D- 6-磷酸果糖的转变.β-D- 6-磷酸果糖继而在相关酶作用下的产物为磷酸烯醇丙酮酸,接而经PK产生丙酮酸.另一方面,乙醇被氧化为乙醛、醋酸后,经ACS合成乙酰辅酶A.有报道表明, 敲除编码G6PI的基因后,鼠细胞中甘油酯含量减少,且油脂代谢中的磷脂酸磷酸酶Ⅰ基因也随之出现表达下调[17]; 在莱茵衣藻中过表达ACS2后,导致叶绿体中碳流向乙酰辅酶A合成的方向富集,从而使得三酰甘油含量提高了2.4倍[18].G6PI、 PK及ACS的表达上调可能通过增加丙酮酸、乙酰辅酶A的含量以增强脂质合成代谢.
在淀粉代谢中,α-D-葡萄糖-1-磷酸为重要的代谢中间物质,磷酸葡萄糖变位酶能将其转化为葡萄糖- 6-磷酸,即丙酮酸的前体.相比HWT藻株,HDLC藻株中内切葡聚糖酶(endo-1, 4-β-D-glucanohydrolase, EGA)、 葡聚糖合酶(glucan synthase, BGS)和果胶酯酶(pectinesterase, PA)均出现表达下调.PA促进ADP-葡萄糖(由α-D-葡萄糖-1-磷酸转化而来)生成直链淀粉; BGS利用α-D-葡萄糖-1-磷酸合成1,3-β-D葡聚糖,继而被转化为D-葡萄糖; EGA则利用以α-D-葡萄糖-1-磷酸为底物合成的纤维素,合成纤维糊精,而后经系列反应产物转变为D-葡萄糖.淀粉代谢途径中3个关键DEGs的下调可能抑制消耗α-D-葡萄糖-1-磷酸的反应过程,间接等导致丙酮酸含量的增加,从而使得碳流定向于脂质合成代谢.
结 语
沉默acs2和cis1基因且过表达DOF转录因子的3基因共调控重定向碳氮代谢中的碳流流向,增强了脂质合成代谢,为莱茵衣藻脂质合成代谢机制研究提供了理论参考,为研究微藻细胞代谢定向调控提供方法.针对以上差异基因富集的代谢通路,后续将对构建的高油脂藻株进行深入的油脂代谢转运和分泌机制的研究.