2.1 生物滞留池中门水平微生物特征分析
2.1.1 微生物多样性和丰富度分析
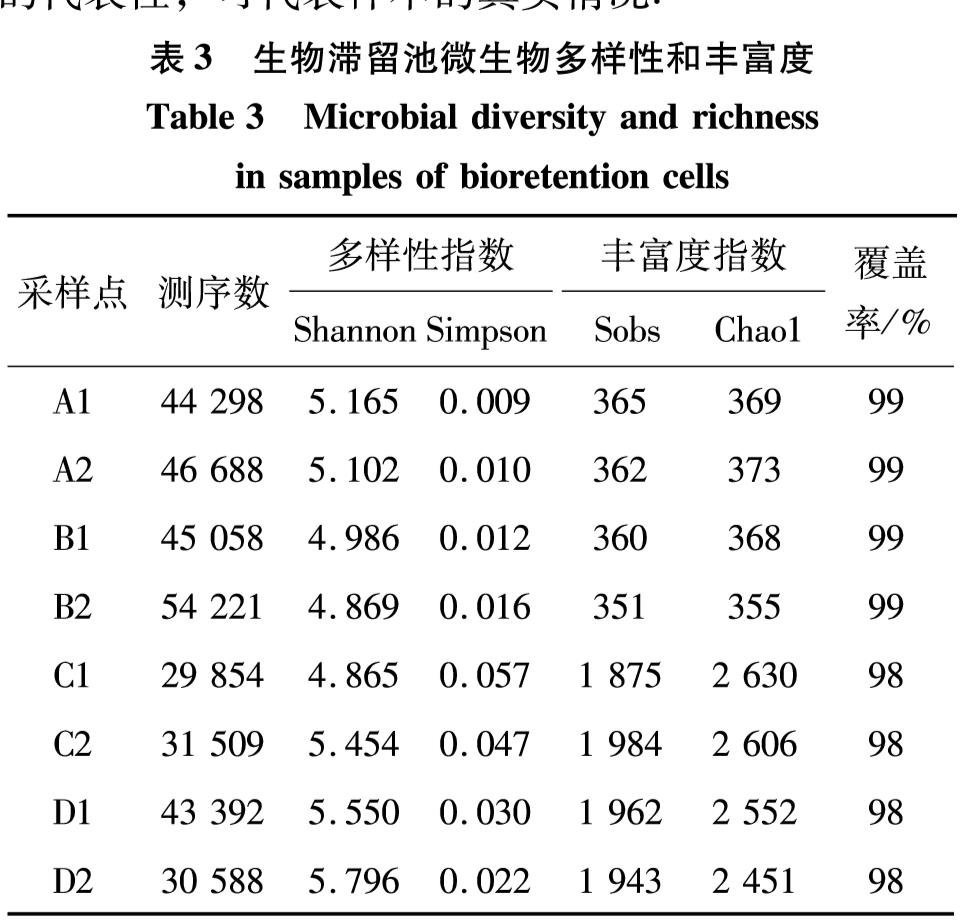
DNA样本的多样性分析可以反映群落的微生物多样性和丰富度,结果如表3.样本文库覆盖率均大于98%,说明测序结果对样本真实情况具有良好的代表性,可代表样本的真实情况.
表3 生物滞留池微生物多样性和丰富度
Table 3 Microbial diversity and richness in samples of bioretention cells
从多样性来看,样本A1、A2、C2、D1和D2的Shannon指数高于样本B1、B2和C1,而样本A1、A2、B1和B2的Simpson指数低于样本C1、C2、D1和D2.一般而言,Shannon指数越高或Simpson指数越低都代表多样性越高.综合考虑这两个指数可知,样本A1多样性最高,即样本A1中的各物种均匀度较好; 样本C1群落多样性最低,结合样本C1的丰富度指数呈现显著高的特点,推测样本C1中可能存在某几种菌为绝对优势菌的情况.A、B组样本的Sobs与Chao1值明显低于C、D组样本的,表明在A、B组样本中物种的丰富度较C、D组更低,这一差异可能与A、B组样本仅稀疏种植单一植物,且总工程土壤厚度较浅有关.
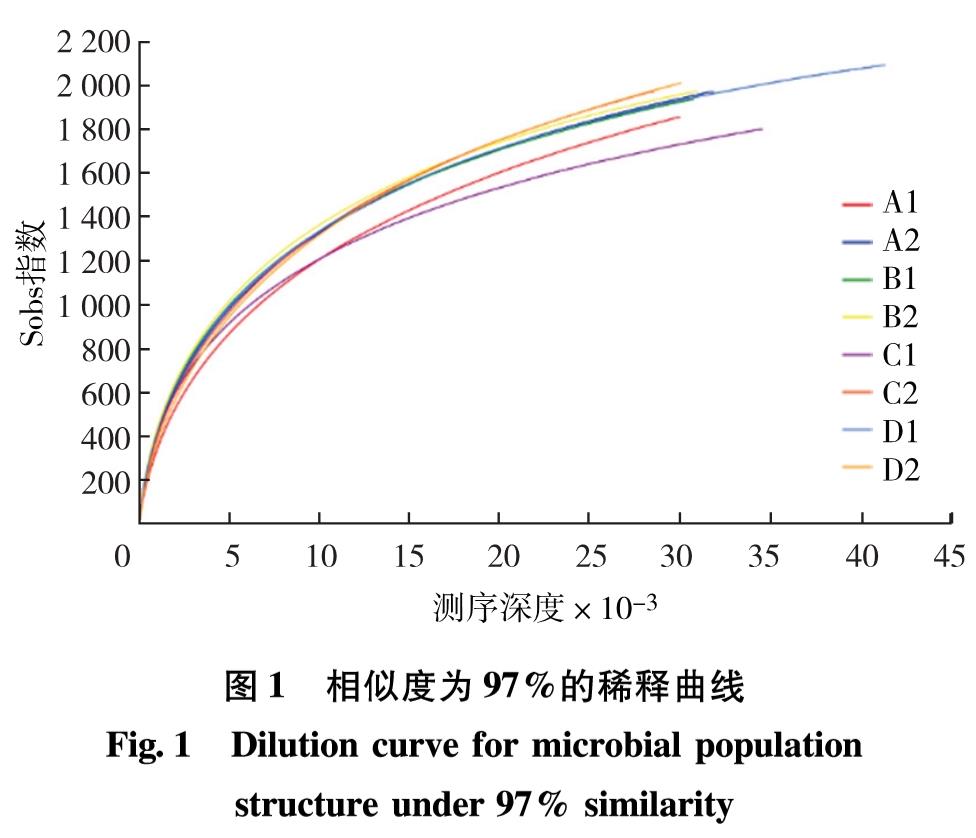
选择97%相似度的OTU,计算不同随机抽样下的Alpha多样性指数,根据多样性指数Sobs(表征实际观测到的物种数目),制作曲线图(图1),即为稀释曲线.观察样本数据的稀释曲线趋向平坦,说明测序数据量合理,更多的数据量只会产生少量的新物种,可初步判定本次测序结果合理可信.
图1 相似度为97%的稀释曲线
Fig.1 Dilution curve for microbial population structure under 97% similarity
图2 样品中门水平细菌(丰度>0.5%)的群落结构及分布
Fig.2 Bacterial community structure and distribution of the samples at phylum level(abundance >0.5%)
2.1.2 生物滞留池中门水平微生物组成差异分析
将相对丰度低于0.5%的门类都合并为others,得到门水平种群分类图,8个土壤样品在门水平上的细菌群落组成如图2所示.由图2可见,8个样品中微生物群落结构组成总体相似,但各种微生物相对丰度略有差异,细菌群落结构在门分类水平上具有较高的多样性,经调查,样品A1、A2、B1、B2、C1、C2、D1和D2中细菌门类检测结果分别为37、37、38、35、34、36、34和35个.
种群分类发现,本研究中8个生物滞留池表层土壤内,相对丰度>0.5%的细菌共有10个门.其中,变形菌门(Proteobacteria)、放线细菌门(Actinobacteria)、拟杆菌门(Bacteroidetes)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)和厚壁菌门(Firmicutes)6个门类是优势细菌菌门,其平均占比依次为29.67%、 21.53%、 12.82%、 12.73%、 10.67%和3.91%,总和占群落的93.49%~94.80%.变形菌门(Proteobacteria)在8个样本中丰度占比最大,样本A1、A2、B1、B2、C1、C2、D1和D2的相对丰度分别为28.69%、29.69%、25.48%、27.55%、33.22%、31.12%、30.70%和30.82%,故变形菌门(Proteobacteria)是优势菌门.有研究发现,变形菌门在生物滞留池[13-14]、人工湿地[15-17]和草地[18]的微生物氮磷去除中起到主要作用,其在生物滞留设施中相对丰度约为25%~68%[7-10].另外,绿弯菌门(Chloroflexi)和厚壁菌门(Firmicutes)对反硝化也有至关重要的作用[16-17].研究发现,样本D1、D2(实验柱对照体系)与样本B1(高污染道路生物滞留池)中绿弯菌门(Chloroflexi)较其他生物滞留池更多,样本C1与C2(建筑小区生物滞留池)中厚壁菌门(Firmicutes)的相对丰度相对与其他6个样本呈显著优势.ZUO等[10]的实验室模拟中,运行良好的实验柱中厚壁菌门(Firmicutes)和绿弯菌门(Chloroflexi)的相对丰度波动范围分别为5%~20%和2%~11%,本研究的结果亦在该范围中.
硝化螺菌门(Nitrospirae)是一类革兰氏阴性菌,其中的硝化螺菌属(Nitrospira)细菌可将亚硝酸盐氧化成硝酸盐,是直接与径流污染处理相关的菌群,硝化螺菌门在样品A1、A2、B1、B2、C1、C2、D1和D2中的相对丰度分别为0.77%、0.67%、1.27%、0.80%、1.26%、1.18%、1.38%和1.58%; 样品D2中硝化螺菌门的相对丰度最大,是样品A1和A2的2倍.
2.2 生物滞留池中属水平功能微生物差异分析
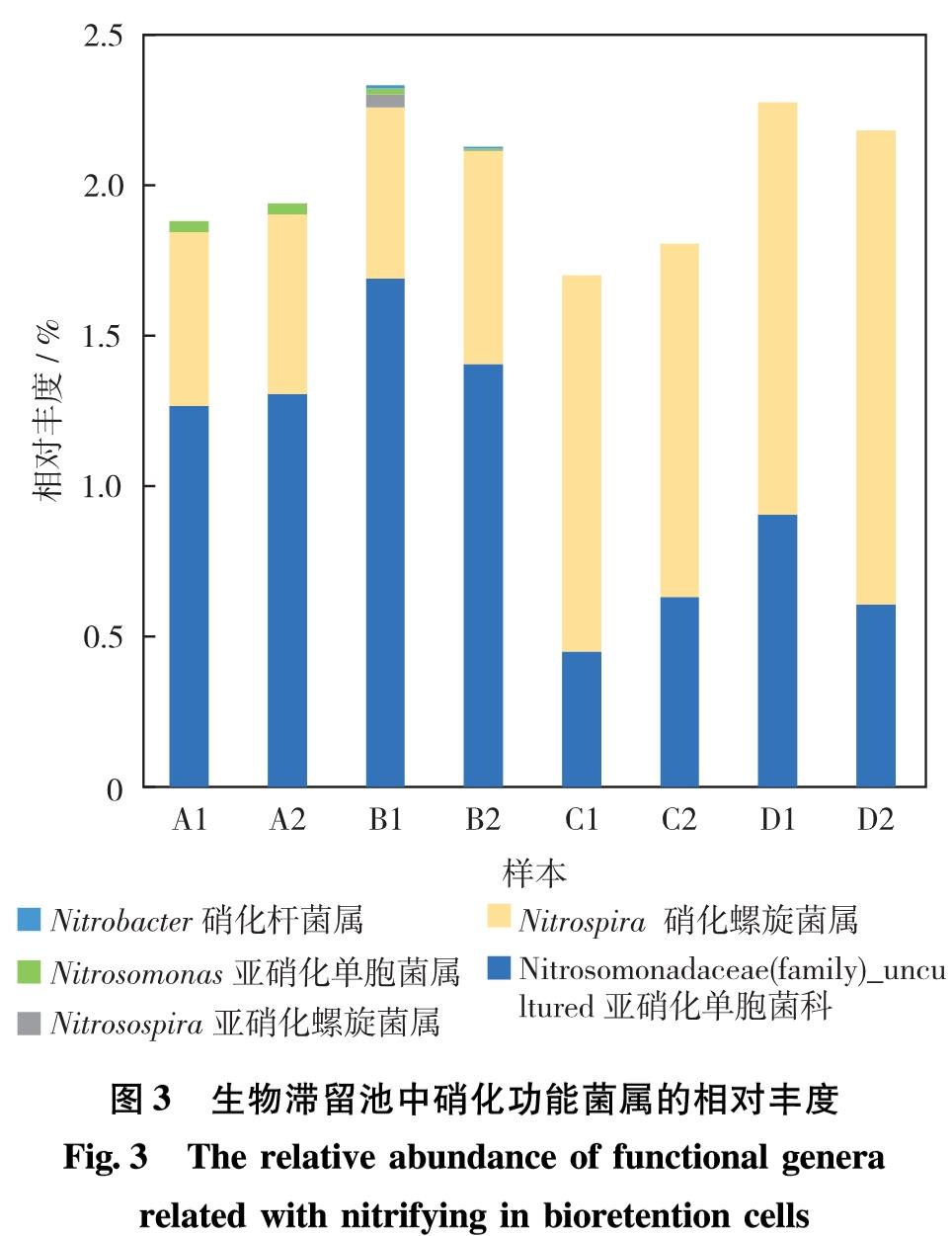
据文献[19-21]研究,硝化杆菌属(Nitrobacter)与硝化螺旋菌属(Nitrospira)的细菌都具有硝化功能; 亚硝化单胞菌属(Nitrosomonas)、 亚硝化螺菌属(Nitrosospira)和亚硝化球菌属(Nitrosococcus)具有氨氧化功能; 不动杆菌属(Acinetobacter)、 假单胞菌属(Pseudomonas)、 芽孢杆菌属(Bacillus)、 硫杆菌(Thiobacillus)、 赖氨酸芽孢杆菌属(Lysinibacillus)、 罗思河小杆菌属(Rhodanobacter)、 热单胞菌属(Thermomonas)和克氏杆菌属(Klebsiella)等具有反硝化的作用,参与氮的转化.因此,重点关注8个典型生物滞留池中相关功能菌属的总相对丰度(图3和图4).调查不同样本中以上几种硝化与反硝化功能菌属的相对丰度,发现硝化细菌与反硝化细菌在不同样本中占比不同.
图3 生物滞留池中硝化功能菌属的相对丰度
Fig.3 The relative abundance of functional genera related with nitrifying in bioretention cells
如图3,在生物滞留池中检出的相对丰度大于0.1%的具有硝化相关作用的微生物主要分布在5个菌属中,包括亚硝化单胞菌科(Nitrosomonadaceae)、硝化螺旋菌属(Nitrospira)、 亚硝化螺旋菌属(Nitrosospira)、 亚硝化单胞菌属(Nitrosomonas)和硝化杆菌属(Nitrobacter). 就硝化细菌的总量而言,各生物滞留池样本相差不大,其相对丰度占所有菌总和的2%左右.其中,亚硝化单胞菌科(Nitrosomonadaceae)属于β-变形菌门(β-proteobacteria),硝化螺旋菌属(Nitrospira)属于硝化螺菌门(Nitrospirae).
对比各组样品中硝化菌的结构组成,发现各组内的2个样本差别较小,但组间表现出了明显区别,A组与B组的硝化种群结构相对一致,C组与D组的种群结构相对一致.在样本A1、A2、B1和B2中,相对丰度最高(分别为1.27%、1.31%、1.70%和1.41%)的硝化功能菌是亚硝化单胞菌科的细菌,是可将NH+4-N氧化成NO-2-N的AOB,且在这些样本中,具有氨氧化作用的AOB功能菌属总丰度大于氧化亚硝酸盐为硝酸盐的亚硝酸氧化菌(nitrite oxidation bacteria, NOB); 而在样本C1、C2、D1和D2中,情况刚好相反,占有绝对优势的功能菌属是硝化螺旋菌属,其丰度分别为1.30%、1.18%、1.38%和1.58%.由于本次取样时间为2月底至3月初,比较生物滞留池A组和B组的结构可发现,由于其具有更浅的工程土壤结构,因此旱季保水性更差,土壤结构具有更高的孔隙率,将有利于好氧的氨氧化细菌生长; 而硝化螺旋菌(Nitrospira)的氮代谢方式则更多样,在缺氧或厌氧的环境中能以氢气作为电子供体把硝酸盐还原为亚硝酸盐[22-23],使Nitrospira在样本C1、C2、D1和D2中的氮代谢竞争中占据巨大优势.
人工湿地是另一种处理非点源污染氮负荷的技术手段,与生物滞留池在结构与功能方面存在一定的相似性,比较对两者内部关键硝化反硝化菌属的研究结果[24]可知,亚硝化单胞菌(Nitrosomonas)在生物滞留池和人工湿地土壤中都是主要优势氨氧化细菌,但是在人工湿地中,亚硝化球菌属(Nitrosococcus)和亚硝化螺菌属(Nitrosospira)也是很常见的优势菌,然而这些菌属在本次生物滞留池的研究中则完全没有发现.可见生物滞留池与人工湿地等类似设施之间微生物结构可能存在差异.
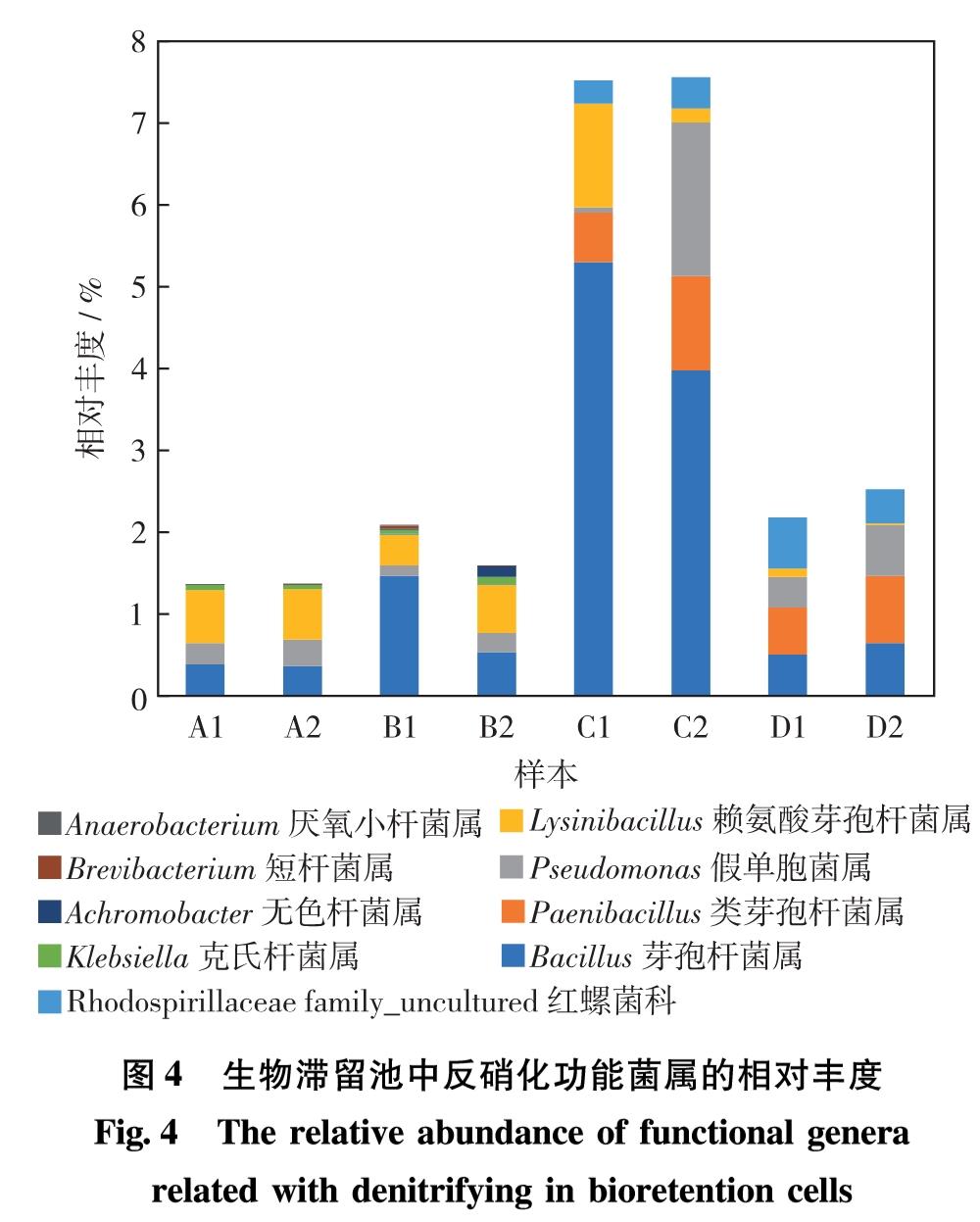
就反硝化菌而言,在生物滞留池中检出的相对丰度大于0.1%,具有反硝化相关作用的微生物主要分布在9个菌属中,各样本中平均相对丰度最高的是芽孢杆菌属(Bacillus), 其次是赖氨酸芽孢杆菌属(Lysinibacillus)与假单胞菌属(Pseudomonas), 如图4.由于硝化与反硝化菌属在自然环境中通常不具有丰度优势,以往研究较少.吴潇[7]的研究注意到,假单胞菌属(Pseudomonas)在生物滞留池的淹没区出现,丰度约为1.39%,与本研究样本C2的丰度水平接近.总体而言,本研究的原位生物滞留池中反硝化生物种类较为丰富,检出的9种反硝化菌属,都是在表面流人工湿地中常见的反硝化功能菌属[17,19].
图4 生物滞留池中反硝化功能菌属的相对丰度
Fig.4 The relative abundance of functional genera related with denitrifying in bioretention cells
样本C1和C2中的反硝化细菌群落的相对丰度占比明显高于其他样本,均超过7.5%,是取自公园和道路4个样点中反硝化细菌相对丰度的3.3~5.1倍.样本C1、C2、D1和D2的生物滞留池都没有50~60 cm的工程土壤,由于反硝化细菌多为厌氧菌,更深的土壤层厚度与植物密集间种,根系丛生带来的湿润土壤环境将有利于在土壤团粒中形成厌氧微环境,因此,具备以上特点的高台生物滞留池样本C1与下沉式生物滞留池样本C2中反硝化功能菌属的相对丰度更高.样本D1和D2的生物滞留池虽然土壤深度较深,但仅有芦苇一种植物,据此推测,生物滞留池种植植物的种类及方式(单种或间种),或将对反硝化细菌的相对丰度产生影响.样本B1和B2中反硝化菌的相对丰度虽然较低,但所检出的反硝化功能菌属的种类丰富度却优于样本C1、C2、D1和D2,这可能与样本B1和B2取自位于非机隔离带中的生物滞留池,接收到的地表冲刷径流中污染物种类更为复杂有关.
2.3 生物滞留池中硝化反硝化功能基因分析
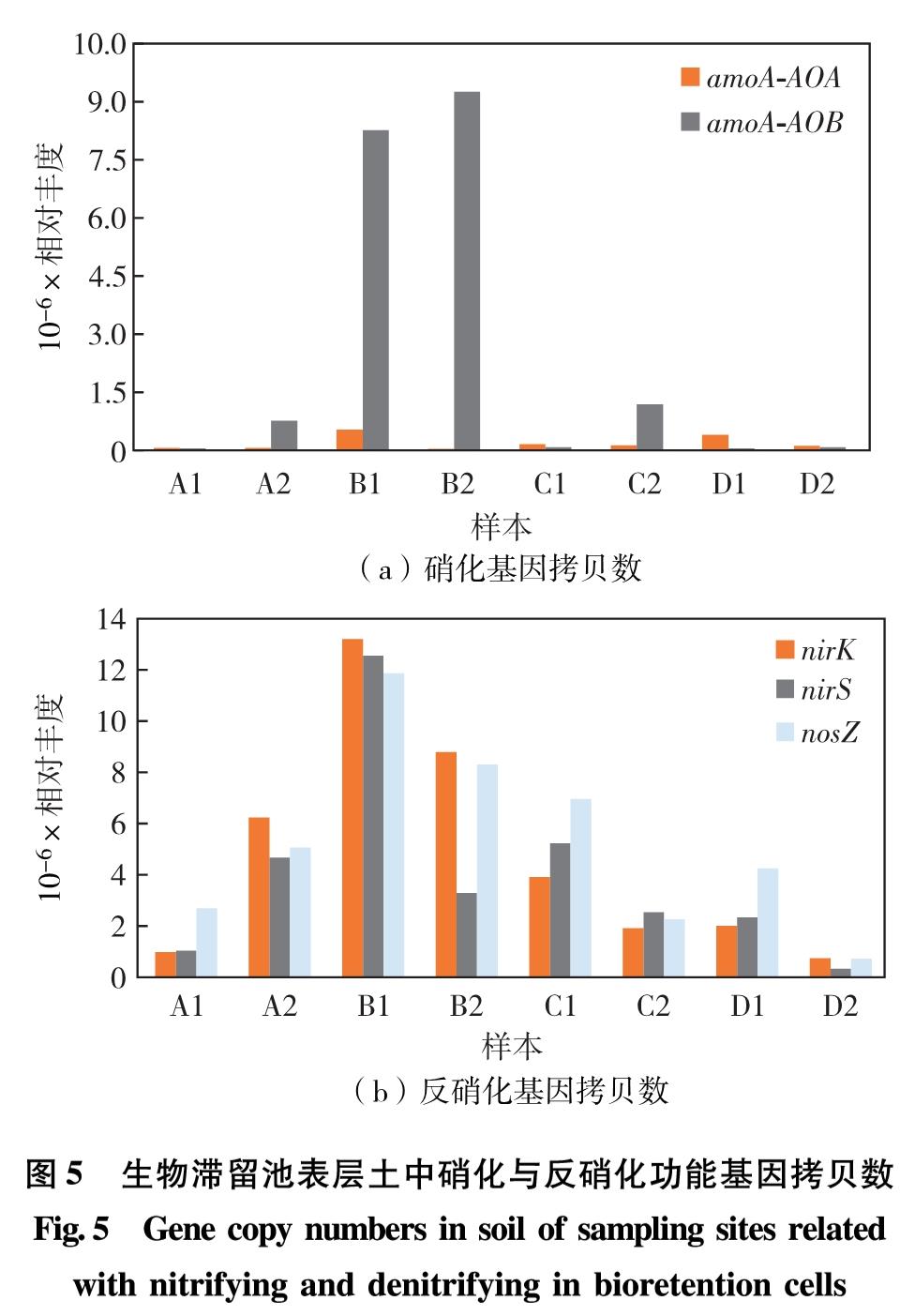
调查样品中表征硝化作用的amoA-AOA与amoA-AOB基因,以及表征反硝化作用的nirK、 nirS和nosZ基因,如图5.研究表明,生物滞留池中硝化反硝化功能基因拷贝数在不同类型的生物滞留池中表现出明显的空间分异.在市政道路类型的生物滞留池样点(B组)中,硝化功能基因丰度与反硝化功能基因丰度与其他样本相比具有显著差异,显著高于其他样本,特别是氨氧化细菌中的amoA基因丰度与其他样本中amoA基因丰度差异超过10倍.样本B1与B2中功能基因的丰度表现出的特异性与其接受的径流污染物浓度较高有关.同时,CHEN等[11]在美国所调查的生物滞留池样本中,处在污水汇流区域、平均淹没时间较其他采样区域更长的位置所测到的反硝化基因nirS、 nirK和nosZ的丰度更高.高污染浓度暴露的长期驯化,使得这些体系中氮转化基因片段的累积更高.
图5 生物滞留池表层土中硝化与反硝化功能基因拷贝数
Fig.5 Gene copy numbers in soil of sampling sites related with nitrifying and denitrifying in bioretention cells
另外,生物滞留池池中监测到的氨氧化细菌amoA基因的平均拷贝数高于氨氧化古菌amoA基因.MORSE等[8,11]的研究也表明,硝化功能基因amoA-AOB的丰度同常较其中的反硝化功能基因nirK、nirS和nosZ的平均丰度低1~3个数量级,这与本研究的调查结果一致.
本研究中发现生物滞留池中amoA-AOB基因的丰度水平最高值约为8.5×106 g-1(每克风干土壤中目的基因的拷贝数),同时也与表面流人工湿地底泥中amoA-AOB基因的丰度水平相近,低值仅为5.9×104 g-1,与中国典型温带草原和草甸草原的丰度水平类似.对于反硝化基因丰度,生物滞留池中nosZ的丰度与中国草甸草原中nosZ的丰度比较类似,较德国半自然草原的丰度更低[18,24].故污染负荷较低的生物滞留池硝化反硝化基因的丰度水平更接近自然生境,如污染负荷较高,则有可能更接近人工湿地沉积物.
2.4 生物滞留池中微生物群落分布与环境因子相关性分析
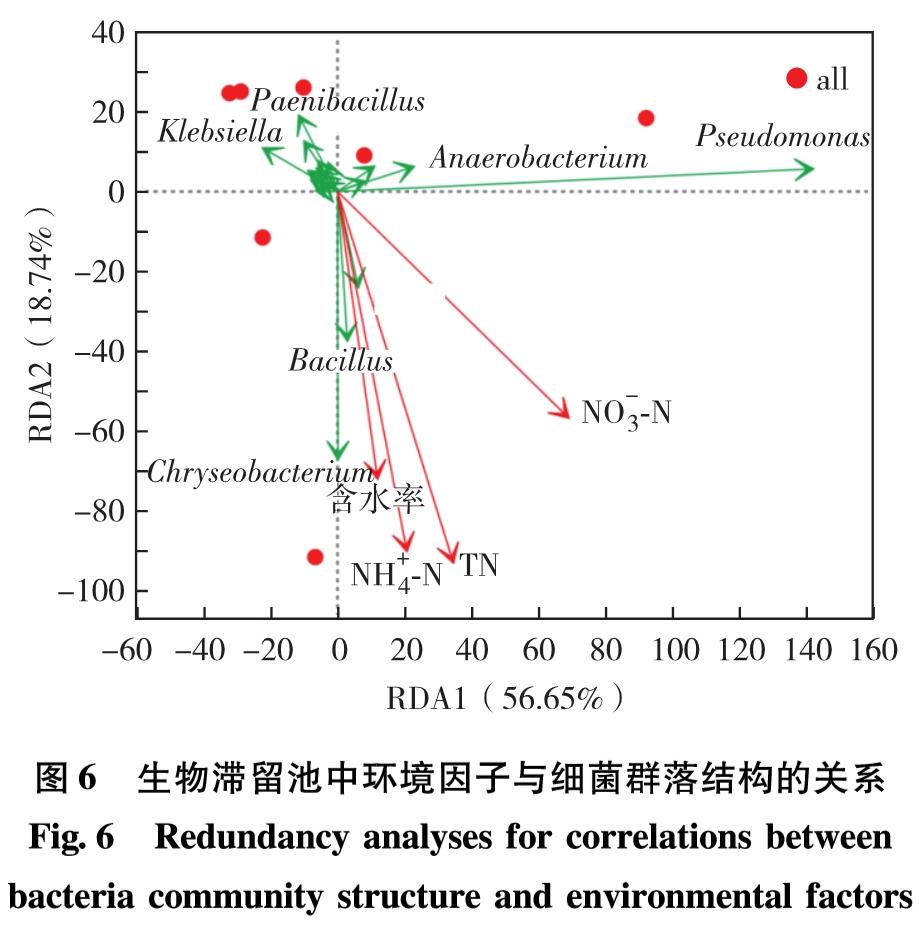
本研究选取NH+4-N质量浓度、NO-3-N质量浓度、总氮(total nitrogen, TN)质量浓度和土壤含水率作为环境因子,同时结合各样本微生物群落结构,选取相对丰度较高的前30种菌属作为样本,通过冗余度分析(redundancy analysis, RDA)研究功能菌属与环境因子的相关性[20],结果见图6.
图6 生物滞留池中环境因子与细菌群落结构的关系
Fig.6 Redundancy analyses for correlations between bacteria community structure and environmental factors
生物滞留池中16S rRNA基因的细菌群落分布与环境因子相关性的RDA分析如图6.坐标轴RDA1和RDA2总解释量为75.39%,相对高的解释量表明了环境因子对生物滞留池中总细菌群落分布影响的可靠性.结果表明,总细菌菌群与生物滞留池表层土中TN浓度(P=0.008, R2=0.963 1)和NH+4-N浓度(P=0.03, R2=0.956 1)显著正相关; 与NO-3-N浓度和土壤含水率呈正相关性但弱相关.解析物种丰度前30的菌属,发现其中受环境因子影响较大的几个菌属中Chryseobacterium、 Paenibacillus、 Bacillus和Pseudomonas等皆为反硝化菌.故判断生物滞留池中反硝化菌更容易受到环境因子的影响.