2.2.1 萌发时间
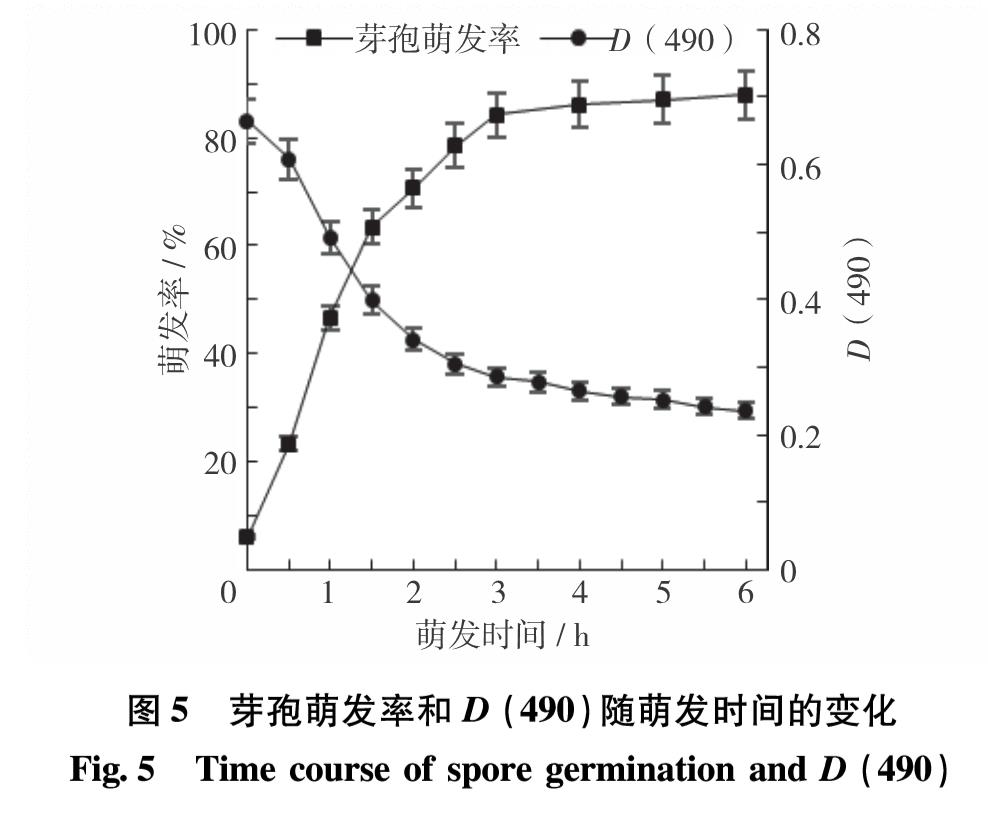
如图5所示,随着萌发时间的增加,D(490)逐渐下降,芽孢萌发率逐渐增加,在4 h时萌发趋于稳定,芽孢萌发率超过85%. 通常芽孢萌发所需的时间较短,且不同菌种的芽孢之间存在差异. SHARMA等[12]研究发现,科氏芽孢杆菌(B. cohnii)芽孢在3 h时,萌发率可达98%,而嗜碱芽孢杆菌(B. pseudofirmus)芽孢在20 h时才开始萌发,经过10 h才萌发完全. 本实验所用的B8芽孢萌发比较迅速,3 ~ 4 h就能萌发完全. 芽孢萌发时间与不同菌种芽孢内膜所含的营养萌发受体量有关,萌发受体量多,芽孢萌发就快,萌发受体量少,芽孢萌发所需时间长,甚至能使芽孢进入深休眠状态[13]. 此外,芽孢萌发速率与所用萌发剂、无机盐、环境pH值和热处理等有关.
图5 芽孢萌发率和D(490)随萌发时间的变化
Fig.5 Time course of spore germination and D(490)
2.2.2 萌发剂
芽孢萌发的萌发剂大多为氨基酸类,本研究对9种萌发剂进行了考察,发现肌苷和L-丙氨酸能较好地促使芽孢萌发,其他几种氨基酸促萌发效果较差(图6). 对肌苷质量浓度作进一步考察发现,在0~0.1 g/L随着质量浓度的增加,芽孢萌发率迅速升高; 在0.1~2.0 g/L,芽孢萌发率增加变缓; 肌苷浓度高于2.0 g/L时,芽孢萌发率几乎不再增加. 可见,适宜的肌苷质量浓度为2.0 g/L. 不同菌种的芽孢所需萌发剂的种类和用量存在差异. 如科氏芽孢杆菌DSM6307(B. cohnii DSM6307)芽孢的最适萌发剂为肌苷,最适浓度为15 mmol/L,浓度过高反而抑制芽孢萌发[14]. GOUNINA-ALLOUANE等[15]发现,利用1~100 mmol/L的L-丙氨酸或0.01~10.00 mmol/L的肌苷能促使蜡状芽孢杆菌(Bacillus cereus)迅速萌发. 除了糖类、氨基酸和嘌呤核苷等营养萌发因子,非营养萌发因子以及高压、热处理等物理因素也能诱导芽孢萌发.
图6 不同萌发剂和肌苷浓度对芽孢萌发的影响
Fig.6 Effects of different germinators and inosine concentration on spore germination
2.2.3 pH值
环境条件是影响芽孢高效萌发的因素之一,如图7,随着pH值的增加,芽孢萌发率呈先增后降的趋势. pH值为10时,芽孢萌发率最高,为86.25%; pH值为9和11时,萌发率分别为74.38%和65.63%,相对较高; pH值低于8或高于12时,芽孢萌发率急剧下降. 因此,芽孢萌发适宜的pH值范围为9~11. 不同菌种的生长环境不同,导致其芽孢萌发所需的pH值也存在差异. VERCAMMEN等[16]研究发现,在800 MPa的高压下、pH值为4的缓冲液能促使凝结芽孢杆菌LMG6326(B. coagulans LMG6326)芽孢萌发. SETLOW等[17]研究表明,pH值为7.3~7.5时,枯草芽孢杆菌(B. subtilis)芽孢开始萌发. 本实验所研究的菌株B8筛选于碱性环境,因此在碱性条件下更有利于该菌株芽孢的萌发.
图7 pH值对芽孢萌发的影响
Fig.7 Effect of pH on spore germination
2.2.4 金属离子
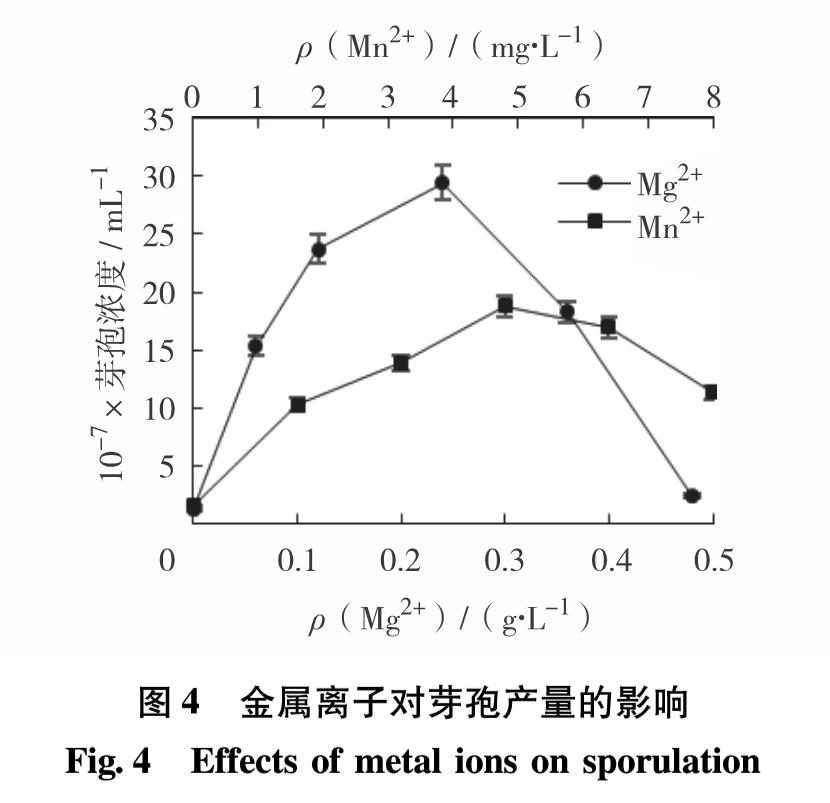
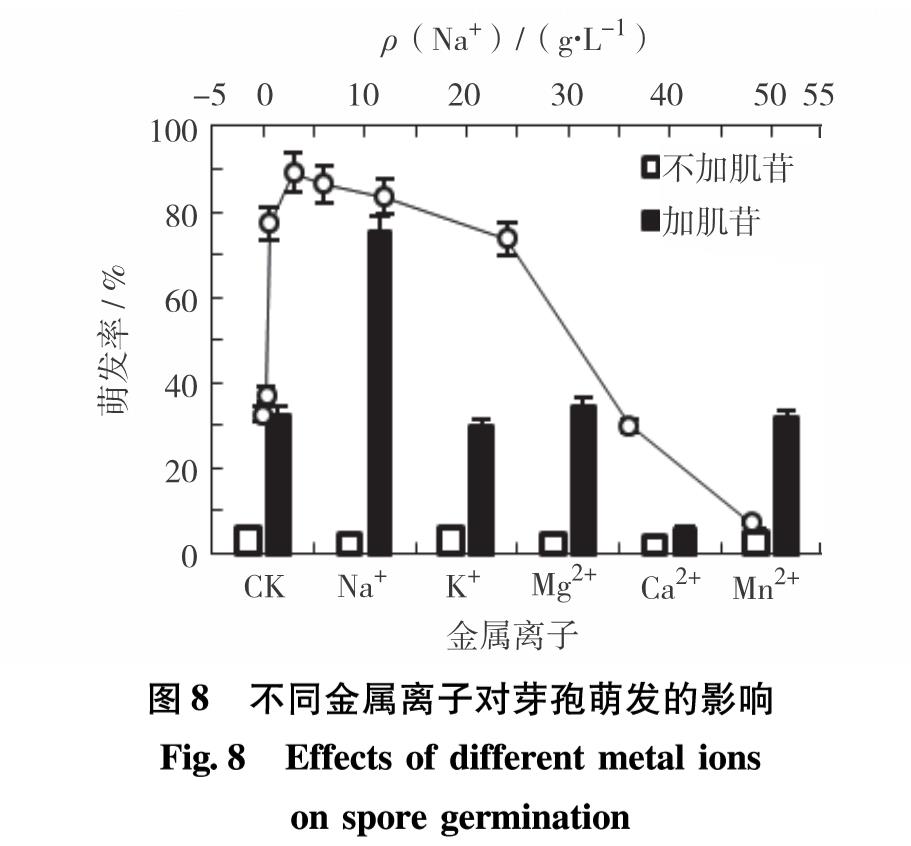
图8显示了不同金属离子对芽孢萌发的影响.由图8可见,不加肌苷的系列中,芽孢几乎不萌发,可见芽孢萌发必须要有萌发剂的参与. 但除肌苷外,一些金属离子也会对萌发过程产生影响. 在加入肌苷的系列中,Na+的存在使芽孢萌发率高达75.42%; 加入K+、Mg2+和Mn2+芽孢萌发率分别为30%、34.67%和32%,与对照组的32.67%相近; 而加入Ca2+芽孢萌发率仅为6%,芽孢几乎不萌发. 可见Na+能促进芽孢萌发,但Ca2+严重抑制芽孢萌发,而K+、Mg2+和Mn2+对芽孢萌发无明显影响.进一步考察肌苷存在时不同Na质量浓度对芽孢萌发的促进作用. 如图8,随着Na+质量浓度的增加,芽孢萌发率呈先增后减的趋势. Na+质量浓度在0~3 g/L时,芽孢萌发率迅速升高; 在3~24 g/L时,芽孢萌发率缓慢下降; 当质量浓度高于24 g/L时,芽孢萌发率迅速下降. 其中,Na+质量浓度为3 g/L时,芽孢萌发率最大,为89.17%; 质量浓度为0.6 g/L和24.0 g/L时,也具有较高的萌发率,分别为77.29%和73.75%. 由此可见,B8芽孢对Na+质量浓度的耐受范围较宽,在0.6~24.0 g/L皆能保持较高的萌发率.
图8 不同金属离子对芽孢萌发的影响
Fig.8 Effects of different metal ions on spore germination
在混凝土内部环境中,Ca2+的含量较高(如普通水泥中,CaO的质量分数高达50%~55%),这将对芽孢萌发造成严重抑制. 芽孢如果不能萌发并生长为营养细胞,就无法诱导碳酸钙的生成,从而大幅降低微生物自修复的效率,因此,必须采取措施解除Ca2+对芽孢萌发的抑制作用. 由图8可知,Na+对芽孢萌发具有明显的促进作用,因此本研究利用Na+的促进作用来抵消Ca2+的抑制作用. 如图9,在含有0.6 g/L的Ca2+体系中加入10.6 g/L的Na+,芽孢萌发率从对照组的6.00%左右上升到30.67%,而加入K+、Mg2+和Mn2+的芽孢萌发率分别为5.33%、4.00%和6.67%,与对照相近,对芽孢萌发无促进作用. 由此可见,Na+确实能在一定程度上解除Ca2+对芽孢萌发的抑制. 从文献调研的情况来看,这是首次证实Na+具有消除Ca2+对芽孢萌发抑制的作用.
图9 含Ca2+体系中,共存金属离子对芽孢萌发的影响
Fig.9 Effects of co-existing metal ions on spore germination in the presence of Ca2+
在确定Na+能有效解除抑制后,进一步考察了不同质量浓度Na+的抑制解除效果. 如图9,在含有0.6 g/L的Ca2+体系中,随着Na+质量浓度增加,芽孢萌发率呈先增后减的趋势. 当Na+质量浓度为24.0 g/L时,芽孢萌发率最高达到82.71%,基本恢复到无Ca2+抑制的水平,表明在此质量浓度时,Ca2+对芽孢萌发的抑制作用被完全解除.
芽孢萌发过程通常伴随有H+和K+等一价阳离子从芽孢内核释放,这可能与芽孢内膜上的离子转运蛋白有关. SOUTHWORTH等[18]研究发现,在蜡状芽孢杆菌(B. cereus)芽孢中存在一种Na+和K+的反向转运蛋白GerN,外界Na+内流促使H+、K+释放,促进芽孢萌发,在其他种属的细菌芽孢中也可能存在类似的离子转运蛋白. SHARMA等[12]研究表明,嗜碱芽孢杆菌(B. pseudofirmus)芽孢萌发需要Na+存在,在萌发体系中加入0.10 mol/L NaCl,芽孢萌发率高达99%,加入0.05 mol/L NaCl,芽孢萌发率为82%,不添加NaCl,芽孢不萌发. 本实验中,Na+能促进B8芽孢的萌发,也可能是芽孢内膜上存在相关离子转运蛋白,通过Na+内流促使内核中其他阳离子外排,伴随着内核吸水,促进萌发.
在芽孢形成过程中,内核生成2,6-吡啶二羧酸(pyridine-2,6-dicarboxylic acid, PDA),会吸收Ca2+,以1:1结合生成螯合物Ca2+-PDA,而芽孢萌发伴随着Ca2+-PDA的释放,当外界存在Ca2+时,会抑制Ca2+-PDA的释放,进而抑制萌发[13]. 本实验表明,当环境中存在Na+时,相关离子转运通道可能被开启,使内核中H+和K+等离子释放的同时可能也导致了Ca2+-PDA释放,引起内核吸水,从而解除Ca2+的抑制作用,最终促使芽孢萌发.