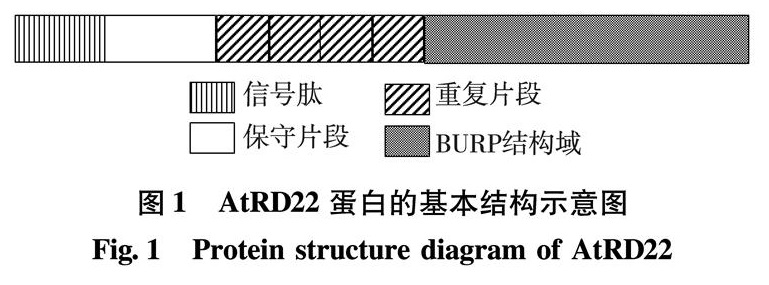
来自拟南芥的一个脱水应答基因(the Arabidopisis thaliana dehydration-responsive gene RD22, AtRD22)的cDNA中有1 179碱基对(base pair, bp)的核苷酸,编码一个含392个氨基酸残基的蛋白,该蛋白属于BURP蛋白家族. BURP蛋白最初是由Hattori等定义的一类在C-端具有保守的BURP结构域的蛋白,其命名取自于4个具有代表性成员:① 油菜花粉粒胚胎发生时表达的一种蛋白(microscope-drived embryo from Brassica napus, BNM2); ② 蚕豆种子中丰度非贮存蛋白(abundant non-storage seed proteins from Vicia faba, USPs); ③ 拟南芥中的一种受干旱诱导的蛋白(dehydration-responsive protein from Arabidopsis thaliana, RD22)[1]; ④ 番茄果实成熟时表达的多聚半乳糖醛酸酶Ⅰ的β亚基(β-subunit of polygalacturonase isozyme I from Lycopersicon esculentum, PG1β)[2].BURP蛋白家族是植物所特有的一类蛋白,已有研究表明,它们在植物的生殖发育、果实成熟以及植物抵抗生物和非生物胁迫中发挥重要功能[3-6].AtRD22蛋白是BURP蛋白家族的典型成员之一,其基本结构如图1.在AtRD22蛋白的BURP结构域中存在较高比例的半胱氨酸、组氨酸和4个保守的半胱氨酸-组氨酸基序,这些氨基酸可能与二硫键的形成有关,也可能具有与过渡金属离子结合的潜力,推测其对蛋白结构的维持和功能的发挥有重要作用,但目前尚未得到验证,因此,通过体外表达获得具有活性的AtRD22蛋白对于研究该蛋白的功能具有重要意义.然而,外源蛋白在大肠杆菌(Escherichia coli, E.coli)中高水平表达时,新生肽链的聚集速率一旦超过蛋白正确折叠的速率就会导致包涵体的形成[7] .如果重组蛋白含有二硫键,而在E.coli体内,由于还原性的环境不利于正确的二硫键的形成,导致重组蛋白链间的错配,也容易导致包涵体的形成.AtRD22蛋白的氨基酸序列中较高比例的半胱氨酸、组氨酸是与二硫键形成有关的氨基酸残基,这可能是该蛋白在E.coli中表达时容易形成包涵体的原因之一,而通过蛋白可溶性分析软件(http://biotech.ou.edu/)预测该蛋白的可溶性为0.因此,如何在体外获得可溶性的AtRD22蛋白是进一步研究该蛋白结构和功能急需解决的技术瓶颈.
图1 AtRD22蛋白的基本结构示意图
Fig.1 Protein structure diagram of AtRD22
融合标签能够在蛋白质的折叠过程中起作用,从而增加重组蛋白质的可溶性表达[8].pET32a载体是一种原核表达载体,利用该载体在E.coli中表达的融合蛋白含有硫氧原还蛋白A(thioredoxin A, TrxA)标签,有研究认为,TrxA具有提高重组蛋白溶解性的能力[9].pSUMO载体是由pET28a改造而来的原核表达载体,它是将一段小泛素相关修饰物(small ubiquitin-related modifier,SUMO)基因插入在pET28a载体的多克隆位点中,编码一个约100个氨基酸残基的小分子泛素样修饰蛋白.利用pSUMO载体在E.coli中表达的融合蛋白含有SUMO融合标签,该标签作为重组蛋白质表达的融合标签和分子伴侣,具有抗蛋白酶水解、提高重组蛋白质可溶性表达等功能[10-11].
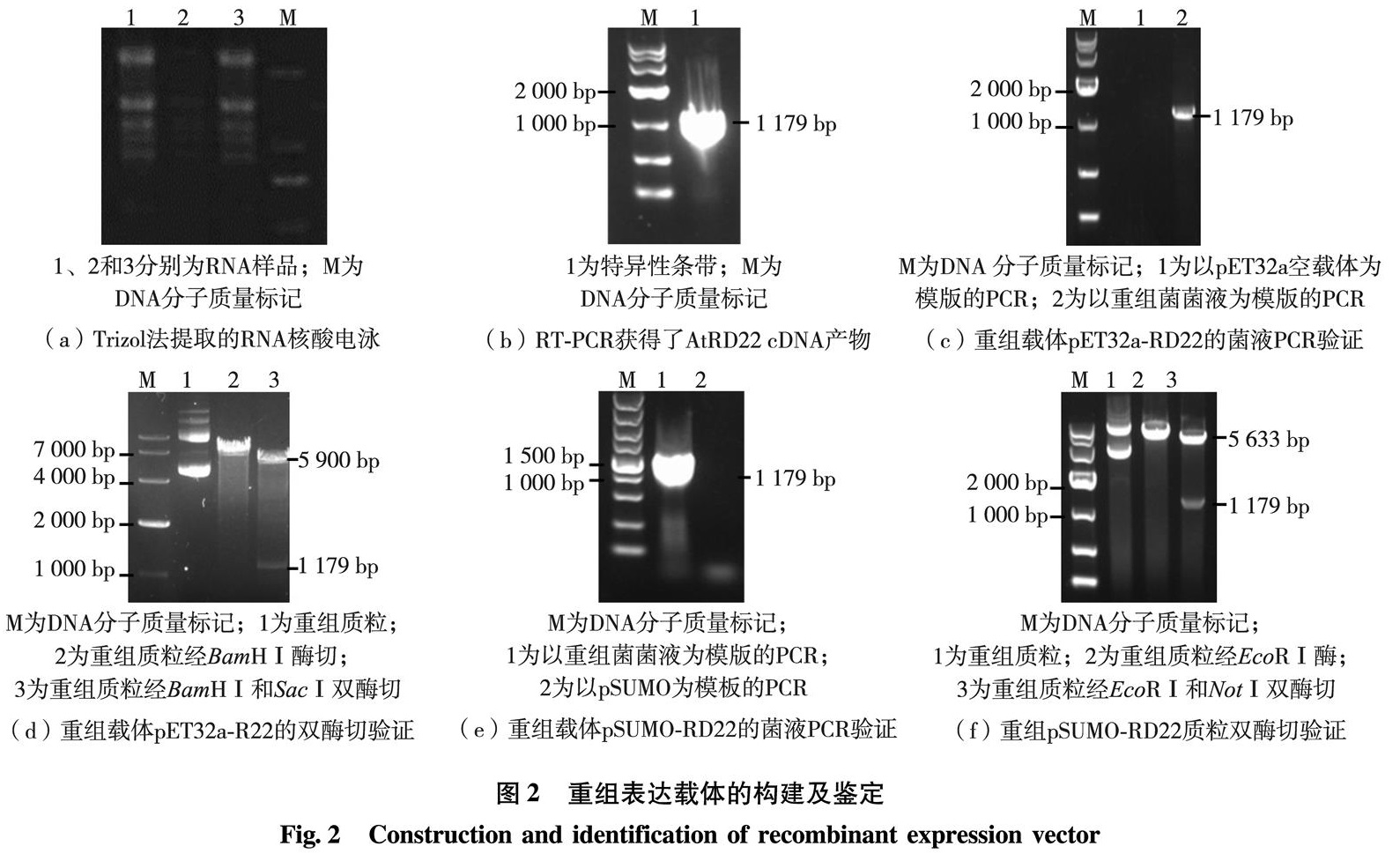
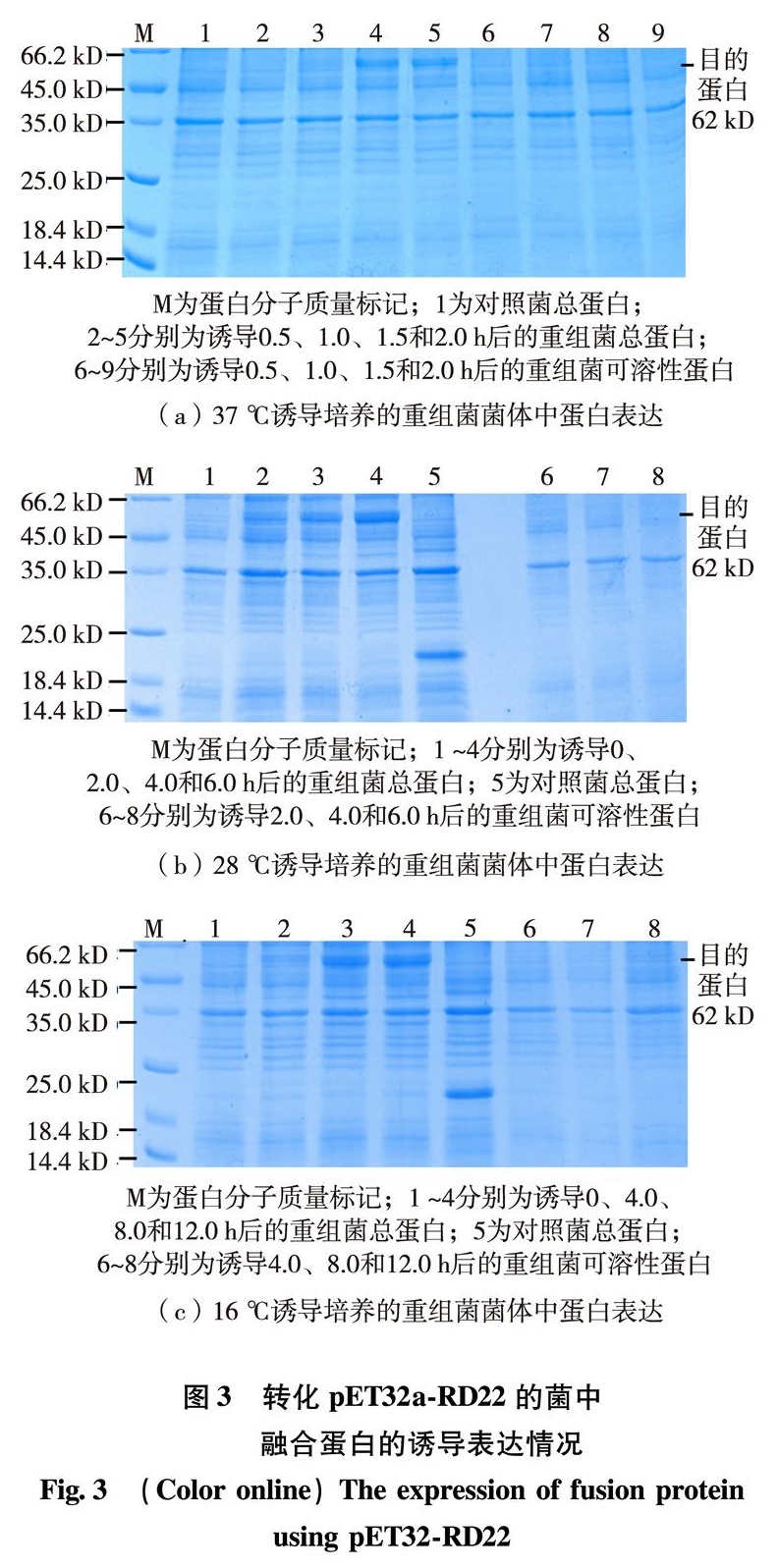
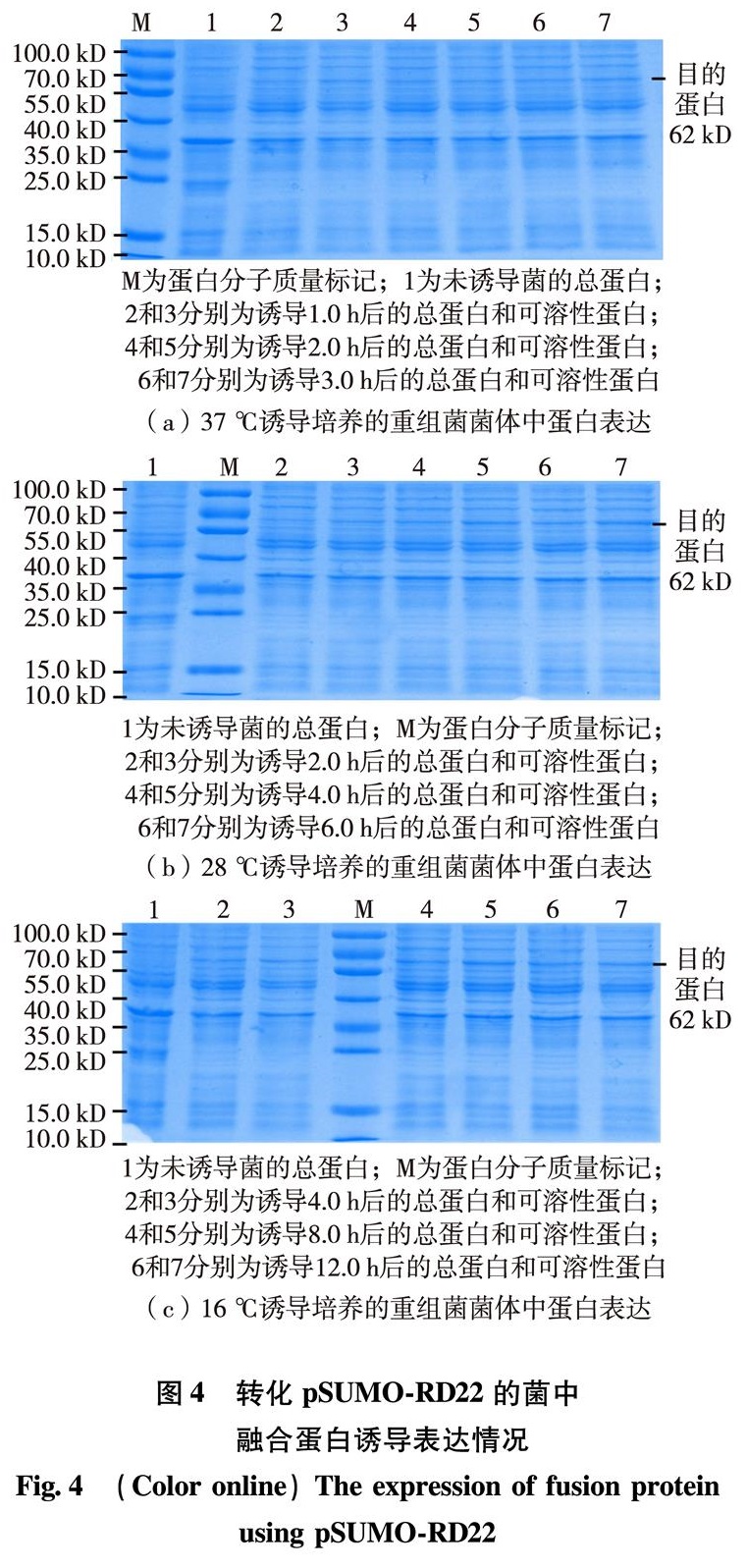
为了获得AtRD22蛋白以研究其结构与功能,本研究利用反转录聚合酶链式反应(reverse transcription-polymerase chain reaction, RT-PCR)的方法先从拟南芥总核糖核酸(ribonucleic acid, RNA)中克隆到了AtRD22全长互补脱氧核糖核酸(complementary deoxyribonucleic acid, cDNA),再分别克隆到原核重组表达载体pET32a-RD22及pSUMO-RD22中,转化E.coli进行蛋白表达.通过一系列条件的优化,找出获得可溶性AtRD22蛋白质的最佳条件并鉴定了融合蛋白的表达.