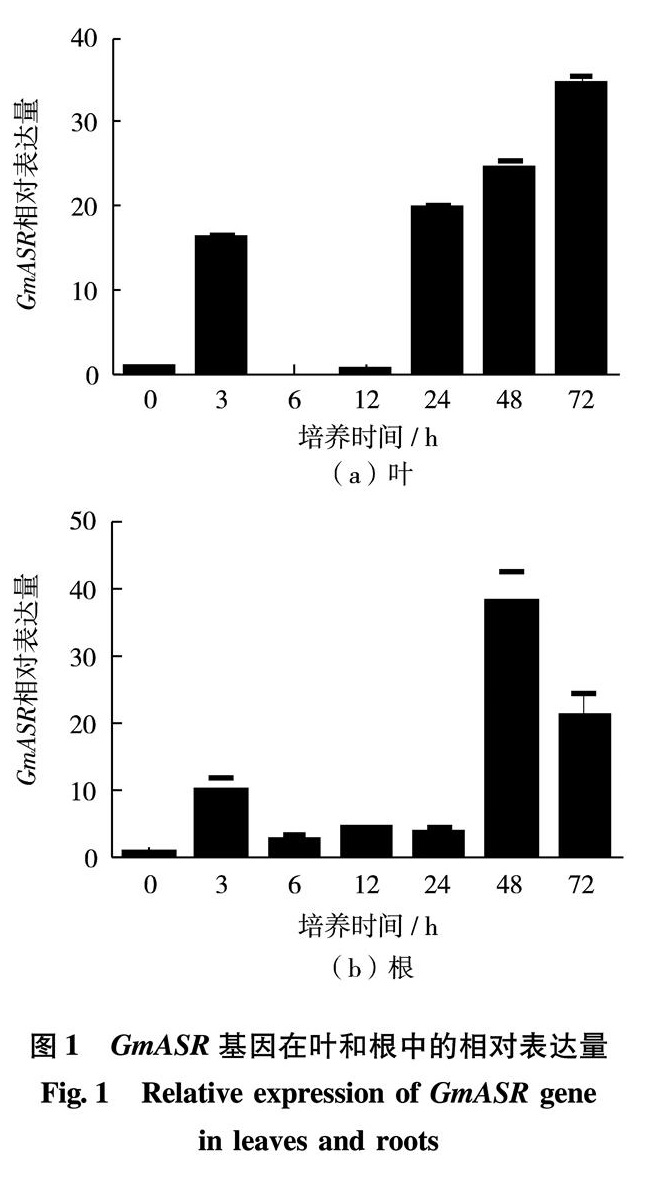
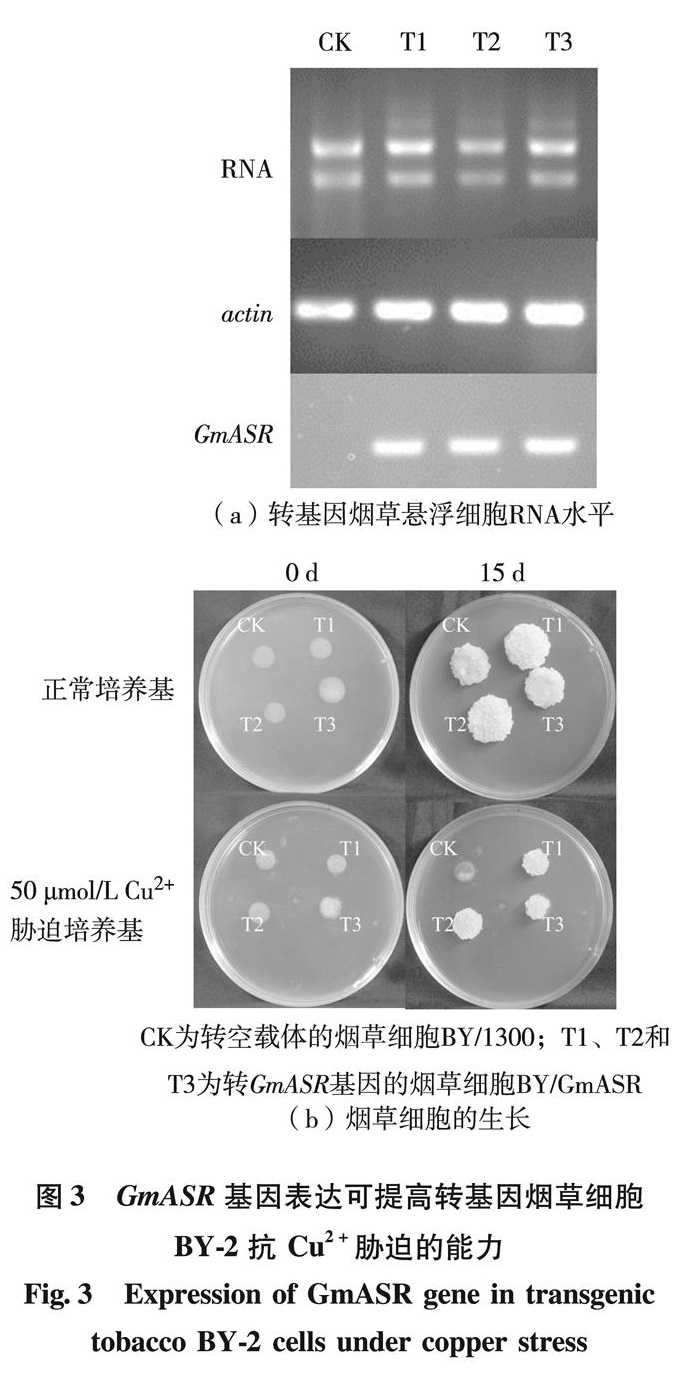
现代冶炼、金属加工、机器制造及其他工业废水中都含有铜,其中,以金属加工和电镀工厂所排废水含铜量最高.这种废水排入水体,会影响水和土壤的质量.Cu2+影响植物生长和发育.植物受到Cu2+胁迫时,会表达多种蛋白质用于减轻胁迫带来的细胞伤害,ASR蛋白是其中重要的一类植物保护性蛋白.本研究结果证明,在Cu2+胁迫过程中,幼苗的叶片和根部GmASR基因的表达量在胁迫3 h和24 h(或48 h)出现2次峰值,表明大豆GmASR基因对Cu2+胁迫存在快速应答和慢速应答.快速应答是植物对周围环境变化的应激性反应,而慢速应答是植物在较长时间胁迫下产生的适应性反应,即胁迫耐受性[19]. GmASR基因的表达受到 Cu2+胁迫的诱导,说明它可能与植物的抗Cu2+胁迫有关.而转基因实验也证明,GmASR基因的过表达确实能提高重组酵母和转基因烟草细胞的抗Cu2+能力.
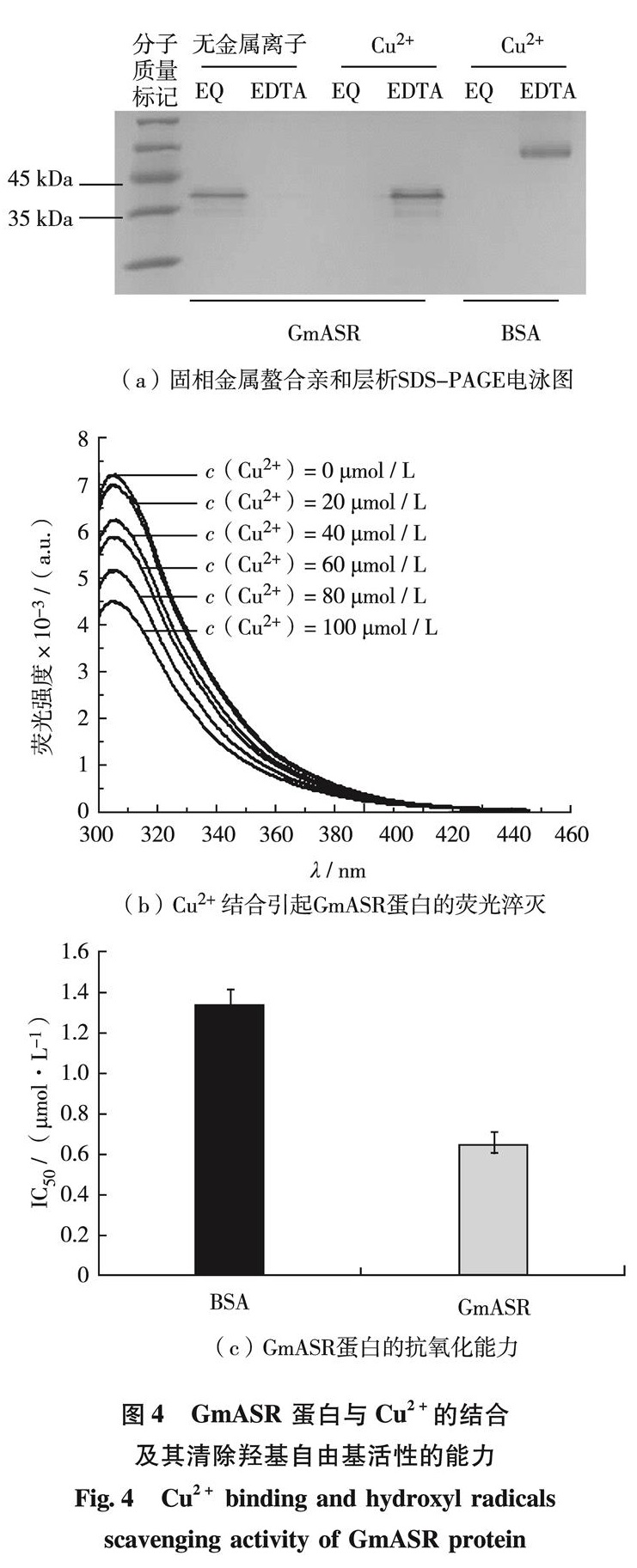
非生物胁迫可以在生理水平和代谢水平上对植物造成影响,如细胞膜稳定性降低、水分丧失、离子平衡失调和离子毒害等,其中,金属(Cu、 Fe、 Cd和Pb等)离子浓度的升高可以催化活性氧物质(reactive oxygen species, ROS)的产生,继而对细胞造成氧化伤害[20].已有研究结果证明蛋白质序列中的组氨酸可结合金属离子,减轻对细胞的氧化伤害.大豆GmPM1和GmPM9蛋白(LEA4)以及柑橘CuCOR19蛋白(LEA2),可通过其序列中的组氨酸结合Fe3+、Cu2+、Ni2+和Zn2+等金属离子,具有清除羟自由基和过氧自由基的功能[21-22].Hara等[18]也证明拟南芥中的AtHIRD11蛋白(LEA2)能减少由Cu2+产生的ROS,并且组氨酸是ROS清除的重要氨基酸.大豆GmASR蛋白含有高比例(9.8%)的组氨酸.文献[15]的研究已证明,GmASR蛋白可以与Fe3+结合,并降低Fe3+催化的羟基自由基.本研究用体外实验证明,GmASR蛋白可直接与Cu2+结合.推测植物受到Cu2+胁迫时,表达的GmASR可螯合细胞内高浓度的Cu2+,减轻Cu2+的毒害作用,降低ROS引起的氧化伤害.因此,利用植物表达ASR蛋白在治理环境中Cu2+污染可能有潜在应用前景.
利用酵母突变体及功能互补实验,可准确地鉴定基因的功能.本实验利用的是Cu2+敏感型酵母ΔCUP2, CUP2基因是Cu2+可诱导的转录因子,当酵母细胞的CUP2基因发生突变后,不能启动下游的Cu2+结合蛋白CUP1的表达,造成细胞内Cu2+积累过多,对细胞产生毒害[23].本研究构建了可表达大豆GmASR蛋白的酵母载体,转化Cu2+敏感型酵母ΔCUP2, 证明GmASR蛋白的表达可以提高酵母突变体耐Cu2+胁迫的能力,即对Cu2+敏感型酵母细胞中CUP2基因的缺失起到了补偿应.这一结果进一步支持了前述“ASR蛋白可通过结合Cu2+,参与植株对Cu2+胁迫的耐受性”的结论.